From environmental input, To physiological output
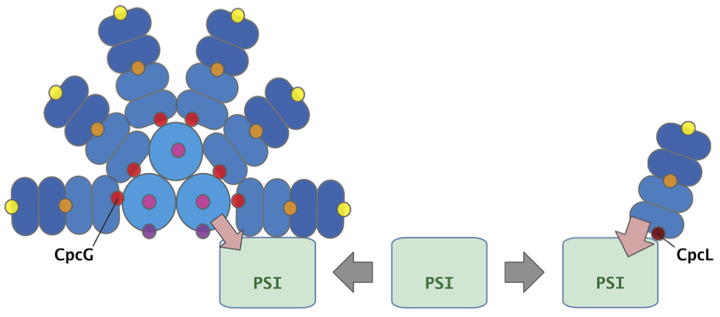
I’m interested in a supermolecular organization of photosystem complexes. My lovely target organisms are cyanobacteria. They perform oxygen-evolving photosynthesis using two photosystems, photosystem I and photosystem II. The name of cyanobacteria comes from their “blue-green” color. But cyanobacteria are more colorful, not only the blue-green but also red, purple, pink and more. The color palette is thanks to their major antenna protein complex “phycobilisome”. Phycobilisome is composed of many colorful chromophore-binding proteins, phycobiliproteins, and their associated linker proteins. The typical phycobilisome consists of several peripheral rods radially projected from a core. A rod-core linker protein CpcG anchors the rod to the core.
Optimization of the photosynthesis is vital for photosynthetic organisms to survive a continually changing environment. Re-organization of light-harvesting antenna systems and photosystems is the primary way for that. Previously, I discovered a novel supercomplex of the photosystem I and a specific phycobilisome, CpcL-phycobilisome, from Anabaena sp. PCC 7120. CpcL-phycobilisome is a rod-type phycobilisome that lacks the core components. It is directly connected with the photosystem I specifically by a linker protein CpcL to form a Phycobilisome-CpcL-Photosystem I supercomplex. Both the typical phycobilisome and the CpcL-phycobilisome can transfer the energy to photosystem I. However, how the cells control the two different types of phycobilisome remains elusive. I aim to reveal the molecular mechanisms of how environmental input can affect the organization of the two distinct phycobilisomes. And also, I want to uncover what is the difference in the physiological roles of the two phycobilisomes as the photosystem I antenna.